Кабанова И.В, Мудрая И.С., Надточий О.Н., Кирпатовский В.И.
Заболевания предстательной железы
(ПЖ) нередко сопровождаются нарушениями функции мочевого пузыря (МП) [1-5]. Можно предположить, что причина сопряженности
заболеваний ПЖ и дисфункции
мочевого пузыря состоит не
только в обструкции, вызываемой увеличенной железой, но в общности регуляторных механизмов, обеспечивающих
кровообращение и иннервацию этих органов. Несмотря на то, что состояние
гладких мышц, ответственных за фазную
сократительную активность и тонус
нижних мочевых путей, является насущным предметом клинических и экспериментальных исследований [5-10], объективные данные о функциональной взаимосвязи МП и ПЖ отсутствуют. Роль автономной нервной системы (АНС) в
регуляции функции МП и ПЖ в настоящее время подтверждается клиническими наблюдениями за эффектами
адренои холинореактивных лечебных
средств и выявлением рецепторов симпатической и парасимпатической систем
в этих органах [5,11-14]. Наши предыдущие исследования показали возможность оценивать активность симпатического и парасимпатического отделов
АНС мочевого пузыря in situ одновременно с состоянием регионарного кровообращения методом гармонического
(спектрального) анализа микровариаций
биоимпеданса отдельного органа [15-17].
Цель настоящего исследования состояла
в оценке функциональной взаимосвязи
мочевого пузыря и предстательной железы в различных условиях накопления
и эвакуации мочи.
МАТЕРИАЛ И МЕТОДЫ
Эксперименты проведены на здоровых половозрелых собаках-самцах (n=7) в
условиях тиопенталового наркоза.
Гармонический анализ биоимпе-данса (ГАБ) для оценки нейрогенной и
сосудистой регуляции МП и ПЖ выполняли с помощью специально разработанного в НИИ урологии программноаппаратного комплекса («Биола», Россия). Оригинальный реограф (импедансный преобразователь), входящий в
состав измерительного комплекса, позволил регистрировать полный (базовый)
биоимпеданс с разрешающей способностью 50 мОм (диапазон 0–1000 Ом) и
его микровариации в диапазоне ±4 Ом
при разрешении 250 мкОм посредством
специальных Ag-AgCl электродов.
Базовый и переменный компоненты импеданса исследуемых органов
регистрировали с помощью двух пар
хлорсеребряных электродов, прикрепленных к стенке МП, а также к правой и
левой долям ПЖ. Одновременно измеряли внутрипузырное (детрузорное)
давление с помощью электроманометра
Capto SP 844 через пункционный катетер
(рис.1). Записи в реальном времени производили на персональном компьютере
для последующего анализа регуляторных
процессов в исследуемых органах во
время естественного наполнения МП и в
разном функциональном состоянии мочевыделительной системы (рис. 2).
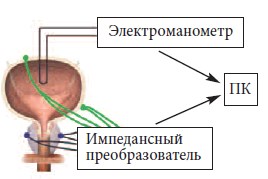
Рис. 1. Схема экспериментальных исследований
МП и ПЖ . ПК – персональный компьютер
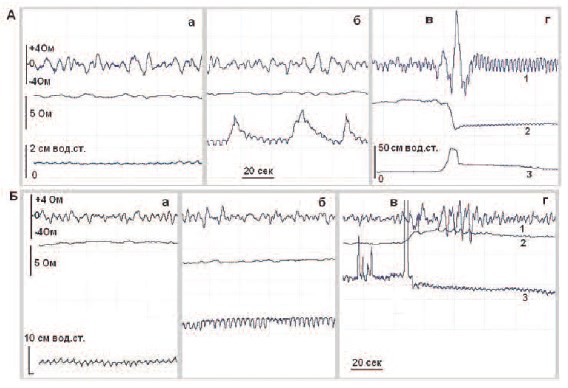
Рис.2. Примеры регистрации переменного (1) и базового (2) импеданса, а также внутрипузырного давления
(3) в МП (А) и ПЖ (Б) у собаки в разные фазы активности МП: арезервуарная фаза, умеренное наполнение, брезервуарная фаза, значительное наполнение, внепосредственно перед мочеиспусканием, гэвакуаторная фаза, мочеиспускание
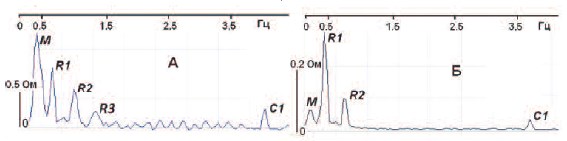
Рис.3. Примеры спектрограмм МП (А) и ПЖ (Б) собаки, полученные Фурье-преобразованием переменных
составляющих регионарных импедансов в резервуарную фазу при значительном наполнении МП. М – пик
Майера, R1 – респираторный пик и его вторая гармоника R2 (в спектре МП присутствует также гармоника
R3), С1 – кардиальный пик
Помимо анализа одновременных
записей импеданса и детрузорного давления во временных координатах, для
оценки нейрогенной и сосудистой регуляции органов анализировали спектро-граммы записей, автоматически вычисляемые методом быстрого преобразования Фурье (рис.3). Для получения амплитудного спектра микровариаций биоимпеданса запись переменной составляющей биоимпеданса МП или ПЖ
фильтровали в частотной полосе 0.05–
15.0 Гц и разбивали на эпохи определенной длительности. На эпохе 25,6 сек
(4096 точек, разрешение 0,04 Гц) получали «обзорный» спектр для анализа
низкочастотных вариаций биоимпеданса
на частотах волны Майера (М) и дыхания (R). Эпоху с длительностью 12,8 сек
(2048 точек, разрешение 0,08 Гц) применяли для анализа сердечных (пульсовых)
гармоник биоимпеданса. Для количественной оценки спектральных пиков
амплитудные спектры нескольких эпох
выбранного фрагмента записи усредняли.
Основываясь на результатах наших
предыдущих исследований [15,17], интенсивность симпатических и парасимпатических влияний оценивали по
спектральным пикам биоимпеданса на частотах, соответственно, 0,08-0,14 (пик
Майера, М) и 0,25-0,40 Гц (респираторный пик, R). Регионарное кровообращение оценивали по первой пульсовой
гармонике кардиального (сердечного) С1
пика на частоте 3,5-4,3 Гц. Расчет спектральных параметров производили в
единицах эффективного сопротивления
[18].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
И ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Длительная регистрация давления
в МП и импедансов МП и ПЖ в течение
постепенного наполнения МП позволила
наблюдать функциональные изменения
обоих органов в реальном времени.
Внутрипузырное давление (рис.2, кривые 3) повышалось по мере наполнения
МП во время резервуарной фазы, демонстрируя небольшие колебания при умеренном наполнении, возрастающие при
значительном наполнении мочевого пузыря, затем оно резко возрастало при сокращении детрузора, и было повышен-ным во время эвакуации мочи. Незначительные колебания давления в фазу накопления мочи, не превышающие нескольких сантиметров водного столба,
можно связать с электрической и сократительной активностью гладкомышечных клеток ПЖ и МП, формирующих
миогенный тонус этих органов. Автономная «микродвигательная» активность дектрузора, способная влиять на
соотношение «давление – объем», т.е. на
податливость МП, зарегистрирована у
мышей во время накопительной фазы
[10]. Тоническую электромиографическю
активность наблюдали в наружном
сфинктере уретры крыс в промежутках
фазной активности [19]. В наших исследованиях метод ГАБ позволил выявить
одновременно с медленными изменениями внутрипузырного давления во
время резервуарной фазы медленные колебания базовых импедансов обоих органов (рис. 4, А и Б кривые 1).
Наблюдаемые медленные изменения регионарных показателей базового
импеданса мы расценили как отражение
тонических изменений в состоянии МП
и ПЖ. Основанием для такой трактовки
результатов послужили следующие соображения. Сопротивление электрическому току, – базовый (полный) импеданс, определяется структурой формирующих его тканей, причем различия базовых импедансов обусловлены особенностями клеток, их пространственным
расположением и внеклеточной жидкостью [20,21]. Помимо постоянных
электрических характеристик, органы
(подобные МП и ПЖ), сформированные
из гладкомышечных элементов или содержащие их, наделены миогенным тонусом [8,10-13,22], который формируется в результате изменяющегося состояния сократительных белков и электрической активности гладкомышечных клеток, в свою очередь, влияющих на электрическую проводимость измеряемой
ткани.
Следовательно, базовый импеданс
МП и ПЖ включает как постоянный
компонент, характеризующий строение
измеряемого органа, так и медленно изменяющийся переменный компонент,
обусловленный изменениями электрических характеристик сокращающихся
мышечных клеток, формирующих или
входящих в состав этого органа. Поэтому,
колебания давления в мочевом пузыре,
которые происходили помимо активных
(фазных) сокращений детрузора и не вызывали мочеиспускание, совместно с
волнообразными изменениями базового
импеданса расценивались нами как показатели тонического напряжения детрузора – его тонуса, обусловленного
активностью гладкомышечной стенки.
Аналогично, об изменениях тонуса ПЖ
мы судили по колебаниям ее базового
импеданса.
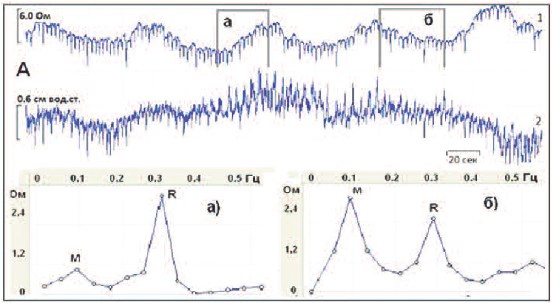
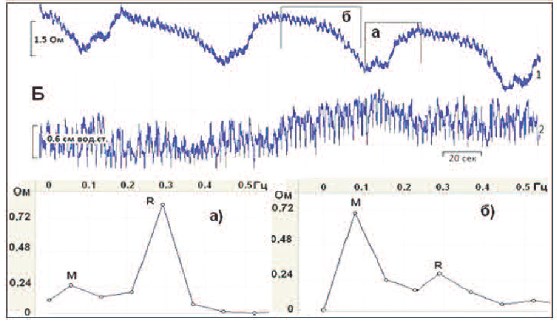
Рис. 4. Короткие фрагменты одновременных записей базового импеданса МП (А,1), ПЖ (Б,1) c детрузорным
давлением (А,2 и Б,2) во время резервуарной фазы со спектрограммами а) и б) соответствующих органов,
построенных для фрагментов записей, обозначенных прямоугольниками аподъема и бснижения
базового импеданса
Периодичность и размах тонических изменений в МП и ПЖ зависели от
фазы мочеиспускания (табл.1). В состоянии покоя, при опорожненном МП, колебания тонуса были незначительными
в обоих органах. По мере наполнения
мочевого пузыря амплитуда колебаний
тонуса в обоих органах и их длительность
увеличивались, причем периодичность тонических колебаний становилась приблизительно одинаковой в МП и ПЖ. Непосредственно перед эвакуаторной фазой
наблюдали противоположные тонические
изменения: в МП колебания тонуса становились чаще, сохраняя высокую амплитуду, а в ПЖ зарегистрированы продолжительные и возросшие троекратно
редкие колебания тонуса.
Наблюдения за колебаниями тонуса МП и ПЖ во время резервуарной
фазы при значительном наполнении
МП, когда тонические изменения в
обоих органах происходили с одинаковой периодичностью, выявили интересные закономерности. Давление и базовый импеданс МП изменялись синфазно, т.е. тонические повышения детрузорного давления часто сопровождались одновременными подъемами базового импеданса (рис. 4, А, а), и, напротив, при снижении тонуса МП,
регистрируемого базовым импедансом,
детрузорное давление имело тенденцию
к снижению (рис. 4, А, б). Такие результаты подтвердили наше мнение, что базовый импеданс МП может служить
показателем его тонуса. Тонус ПЖ, судя
по показателю ее базового импеданса,
также колебался во время резервуарной
фазы, однако связь этого тонуса и детрузорного давления была реципрокной.
Действительно, тонус ПЖ возрастал
(рис. 4, Б, а) в то время, когда давление в
МП снижалось. И, напротив, тонус ПЖ
снижался (рис. 4, Б, б) во время повышения детрузорного давления.
Учитывая, что МП и ПЖ находятся
под постоянным контролем АНС, мы
проанализировали изменения спектральных параметров, характеризующих активность симпатического и парасимпатического звеньев вегетативной регуляции,
происходящих во время повышения или
снижения тонуса этих органов. Наблюдения за базовым импедансом МП и ПЖ показали, что тонические колебания
сопряжены с периодическими изменениями активности обоих звеньев АНС,
судя по изменениям Ми Rпиков. Оказалось, что повышение тонуса как в МП,
так и в ПЖ, было связано с возрастанием
регионарной парасимпатической активности, судя по увеличенному R-пику (а)на рис.4, А и Б) по сравнению с М-пиком,
отражающим симпатическую активность.
Противоположные изменения, а именно,
увеличенный М-пик (б)на рис.4, А и Б)
сравнительно с R-пиком, были зарегистрированы при снижении тонуса обоих
органов, – уменьшении их базовых импедансов.
Таблица 1. Изменения тонуса мочевого пузыря и предстательной железы у
собак, выявленные методом ГАБ в разные фазы активности мочевого пузыря
| Фаза активности МП |
Амплитуда
тонических
колебаний
импеданса МП |
Периодичность
тонических
колебаний
в МП |
Амплитуда
тонических
колебаний
импеданса ПЖ
| Амплитуда
тонических
колебаний
импеданса ПЖ |
Пустой МП
(после мочеиспускания) |
2,82 Ом |
11 сек. |
2,55 Ом |
19 сек. |
Резервуарная фаза 1:
умеренного наполнения МП |
5,85 Ом |
42 сек. |
3,03 Ом |
52 сек. |
| Резервуарная фаза 2: значительного наполнения МП |
8,07 Ом |
74 сек. |
3,36 Ом |
75 сек. |
| Мочеиспускание |
6,06 Ом |
17 сек. |
9,9 Ом |
257 сек. |
Тонус органов мочеполовой системы занимает важное место в механизме функционирования нижних мочевых путей, что отражено в многочисленных клинических и экспериментальных
исследованиях в урологии [9, 11, 12, 22].
Тонус органов мочеполовой системы помимо пассивных элементов, входящих в
состав каждого органа, обеспечивается
электрической и сократительной активностью гладкомышечных элементов,
входящих в их состав. Спонтанный электрический ритм выявлен на протяжении
мочевого тракта, однако его клеточное
происхождение и связь с сократительной
и тонической функцией отдельных органов, а также с их иннервацией до настоящего времени остается предметом исследований [6, 7, 10]. До сих пор роль регуляторных влияний АНС на функциональную активность разных органов
обсуждается с позиций рецепторного аппарата клеточной мембраны гладкомышечных, секреторных и эпителиальных
клеток [4, 11-14].
Полученные нами объективные
данные об одновременных и противофазных изменениях тонуса МП и ПЖ,
частота и размах которых зависят от величины и колебаний давления в МП, а
также выявленные синхронные с колебаниями тонуса изменения Ми Rпиков
спектрограмм этих органов, характеризующих реципрокные отношения симпатического и парасимпатического отделов
АНС, свидетельствует о нейрогенной
природе происходящих тонических изменений. Проведенные исследования методом ГАБ продемонстрировали, что
базовый импеданс может нести информацию о тонусе МП и ПЖ во время резервуарной фазы, а вариации биоимпеданса способны отразить колебания,
вызванные собственной биоэлектрической активностью стенки, которая может
быть связана с нервными влияниями
разных отделов АНС на клетки органов
и кровеносные сосуды.
Метод ГАБ позволил также оценить регионарные регуляторные процессы в МП и ПЖ здоровых собак,
осуществляемые симпатическим и парасимпатическим отделами АНС, и их изменения во время фазной (сократительной) активности МП (рис. 5). Анализ
спектральных параметров, характеризующих активность обоих звеньев нейрогенной регуляции МП и ПЖ, а также
их регионарную сосудистую активность,
выполнен для участков записей перемен-ного импеданса МП, соответствующих
разным фазам функционирования нижних мочевых путей.
Изменения показателей нейрогенной активности в фазу накопления и эвакуации мочи в мочевом пузыре собак
были аналогичными изменениям, наблюдаемым в предыдущих исследованиях на
крысах [15, 17]. Так, во время резервуарной фазы МП пик Майера у собак возрастал на 29-43%, соответственно при
умеренном и значительном наполнении,
что сопровождалось одновременным
снижением на 34-53% R-пика. Наблюдаемые противоположные изменения спектральных Ми Rпиков демонстрируют
реципрокные отношения симпатического и парасимпатического отделов
АНС: активизации симпатической активности в фазу накопления мочи, способствующей снижению тонуса и поддержанию низкого внутрипузырного
давления, и активизации парасимпатической нервной активности в фазу изгнания мочи, ответственной за сократительную функцию детрузора, что соответствует современному пониманию
особенностей функционирования нижних мочевых путей [14].
Аналогичный анализ записей давления в реальном времени и спектральных параметров переменного импеданса
выполнен для ПЖ в разные фазы мочеиспускания (рис. 5, Б). В ПЖ, как и в МП
здоровых собак, выявлена противоположная динамика Ми Rпиков в разные
фазы активности МП, демонстрируя реципрокные отношения симпатической и
парасимпатической иннервации. При
этом изменения спектральных характеристик в ПЖ в фазу накопления и изгнания мочи были противоположными
изменениям этих параметров в МП. Так,
на протяжении резервуарной фазы при
нарастании давления в МП наблюдали
снижение М-пика (на 20-32%) и увеличение R-пика (24-54%), а во время эвакуаторной фазы МП при высоком детрузорном давлении R-пик в спектрограммах
ПЖ уменьшался (на 31%).
Изменения С-пика в обоих органах
происходили параллельно с изменениями М-пика, и в ПЖ также наблюдалась
обратная динамика этого показателя в
сравнении с его изменениями в МП. При
наполнении МП до физиологического
объема кардиальный пик в спектрограммах МП увеличивался (на 36-53% при
умеренном и значительном наполнении),
свидетельствуя об усилении вазоактивности и интраорганного кровотока,
тогда как в ПЖ зарегистрировано равномерное уменьшение Спика по мере наполнения МП.
Таким образом, метод ГАМБ позволил продемонстрировать функциональную взаимосвязь нейрогенных регуляторных процессов, осуществляемых
АНС в МП и ПЖ как при фазных сокращениях детрузора, вызывающих мочеиспускание, так и при тонических колебаниях давления в МП, т.е. при колебаниях
тонуса МП, – небольших сокращениях,
не сопровождающихся мочеиспусканием. При этом роль внутрипузырного
давления явилась определяющей для изменений регуляторных процессов в ПЖ,
что указывало на важность этого показателя, как регуляторного фактора в патогенезе заболеваний ПЖ.
Высокое детрузорное давление во
время эвакуаторной фазы МП (рис. 5, Б,
IV) было сопряжено с преобладанием
симпатической активности ПЖ (рост
пика Майера), причем в это время в МП
активизировалась парасимпатическая
активность (рис. 5, А, IV). Аналогично,
при тонических колебаниях детрузорного давления, усиление симпатической
активности ПЖ и уменьшение парасимпатических влияний зарегистрировано при
высоком детрузорном давлении, когда
тонус ПЖ снижался (рис. 4, Б, б), а тонус
МП возрастал (см. рис. 4, А, а), что сопровождалось усилением парасимпатической активности. Повышение тонуса
предстательной железы и усиление ее парасимпатической активности, напротив,
происходило при тоническом снижении
детрузорного давления (рис. 4. Б, а) в то время как в МП усиливались симпатическая активность (рис. 4, А, б), а
перед мочеиспусканием регионарный
симпато-парасимпатический баланс ПЖ
смещался в сторону парасимпатической
активности (рис. 5, Б, III), тогда как в МП
происходило расслабление детрузора и
преобладали симпатические влияния
(рис. 5, А, III).
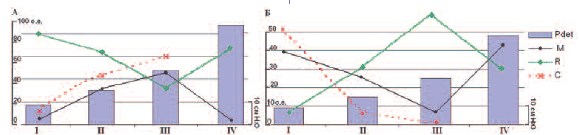
Рис. 5. Относительные изменения спектральных параметров, отражающих регионарную нейрогенную и
сосудистую активность МП (А) и ПЖ (Б) собаки при I –опорожненном МП; II – умеренно наполненном МП;
III – значительно наполненном МП; IV– в фазу эвакуации мочи. Обозначение графиков: Pdet – детрузорное
давление; M пик Майера; R –респираторный пик; С кардиальный пик.
Роль парасимпатической нервной
системы, стимулирующей сократительную функцию детрузора МП, подтверждена в настоящее время многими
клиническими фармакологическими и
экспериментальными in vitro исследованиями [3-6, 14, 23, 24]. На основании исследований иннервации ПЖ и состояния
рецепторов АНС показано, что парасимпатическая нервная система оказывает
модулирующее влияние на секреторную
и тоническую активность ПЖ, а также
увеличивает ее гладкомышечный тонус
[23, 25]. В проведенных нами исследованиях метод гармонического анализа мик-ровариаций биоимпеданса МП и ПЖ
позволил in situ продемонстрировать
роль симпатического и парасимпатического звеньев АНС в регуляции функции
исследуемых органов в разные фазы мочеиспускания.
Можно предположить, что противофазные изменения тонуса МП и ПЖ
указывают на функциональные взаимосвязи этих органов в осуществлении
функции изгнания мочи, когда в ответ на
сокращение детрузора происходит расслабление ткани ПЖ, уменьшая тем
самым сопротивление уретры току мочи.
Ухудшение состояния кровообращения в
ПЖ, наблюдаемое при повышенном
внутрипузырном давлении, может указывать на важность этого патогенетического фактора заболеваний ПЖ при
инфравезкальной обструкции разной
этиологии, а снижение парасимпатической активности ПЖ, которое происходит при активизации сокращений
детрузора, повышении давления и уве-личении его парасимпатической активности, может иметь значение для исследований секреторной функции ПЖ.
В целом, проведенные исследования
свидетельствуют о координирующей роли
АНС в регуляции фазной активности и
поддержании тонуса МП и ПЖ. Выявление существующей функциональной взаимосвязи этих органов у здоровых животных объясняет возможную сопряженность нарушений регуляции этих органов
при заболеваниях каждого из них, которые могут быть как результатом причинно-следственных отношений, – обструкции оттоку мочи увеличенной ПЖ,
так и следствием взаимодействия измененной сократительной функции, нервной
и сосудистой активности МП на регуляторные процессы в ПЖ. Исследование регуляторных процессов, осуществляемых
АНС, методом ГАБ может быть полезным
для диагностики различных функциональных расстройств нижних мочевых
путей и их лечения.
ЛИТЕРАТУРА
1. Вишневский А.Е. Нарушение энергетического метаболизма в патогенезе расстройств мочеиспускания у больных доброкачественной гиперплазией простаты и обоснование их медикаментозного лечения: Дисс. докт.
мед. наук. М., 2007.
2. Тюзиков И.А., Мартов А.Г., Калинченко С.Ю. Новые системные механизмы патогенеза симптомов нижних
мочевых путей у мужчин (литературный обзор). // Бюллетень сибирской медицины. 2012. N 2. С.93-100. 2012.
3. McVary KT, Rademaker A, Lloyd GL, Gann P. Autonomic
nervous system overactivity in men with lower urinary tract
symptoms secondary to benign prostatic hyperplasia// J
Urol. 2005. Vol. 174. P. 1327-1433.
4. Strittmatter F, Gratzke C, Stief CG, Hedlund P. Current
pharmacological treatment options for male lower urinary
tract symptoms. // Expert Opin Pharmacother. 2013.
Vol. 14, N 8. P. 1043-1054.
5. Giannitsas K, Athanasopoulos A. Current understanding
of the interplay between OAB&BPH. // Curr Bladder Dysfunct
Reports, 2013. DOI 10.1007/s11884-013-0178-7
6. Кирпатовский В.И., Фролова Е.В., Надточий О.Н.
Спонтанная ритмическая активность органов мочевой
системы: роль интерстициальных клеток, биологическая
значимость, патофизиологические аспекты. // Экспериментальная и клиническая урология. 2012. N 2.
C. 70-77.
7. McHale NG, Hollywood MA, Sergeant GP, Shafei M,
Thornbury KT. Organization and function of ICC in the urinary
tract. // J Physio. 2006. Vol. 576, Pt 3. P. 689-694.
8. Lin G. Modulation of smooth muscle tonus in the lower
urinary tract: Interplay of MLCK and MLCP. // BJU Int. 2011.
Vol. 108, Pt 2. P. 66–70.
9. Ückert S. Oelke M, Phosphodiesterase (PDE) inhibitors
in the treatment of lower urinary tract dysfunction// Brit J
Clin Pharmac. Special Issue: Uropharmacology Themed
Section. 2011. Vol. 72, N 2. P. 197–204.
10. Smith PP, DeAngelis AM, Kuchel GA. Evidence of central
modulation of bladder compliance during filling phase//
Neurourol Urodynamics. 2012. Vol. 31, N 1. P. 30–35.
11. Ventura S, Pennefather JN, Mitchelson F. Cholinergic
innervation and function in the prostate gland. // Pharmac
Therapeut. 2002. Vol. 94, N 1–2. P. 93–112.
12. Witte LP1, Chapple CR, de la Rosette JJ, Michel MC
Cholinergic innervation and muscarinic receptors in the
human prostate. // Eur Urol. 2008. Vol.54, N 2.
P. 326–334.
13. Michel MC, Sand C. Effect of pre-contraction on alfaadrenoceptormediated relaxation of rat urinary bladder. //
World J Urol. 2009. Vol. 27. P. 711–715.
14. Ochodnicky P, Uvelius B, Andersson KE, Michel MC.
Autonomic nervous control of the urinary bladder. // Acta
Physiol (Oxf). 2013. Vol. 207, N 1. P. 16-33
15. Мудрая И.С., Ибрагимов А.Р., Кирпатовский В.И., Ревенко С.В., Нестеров А.В., Гаврилов И.Ю. Оценка функционального состояния мочевого пузыря крыс методом
Фурье-импедансной цистометрии. // Экспериментальная и клиническая урология. 2010. N 3. C. 21-26.
16. Нестеров А.В., Гаврилов И.Ю., Селектор Л.Я., Мудрая
И.С., Ревенко С.В. Фурье-анализ вариаций биоимпеданса
пальца человека// Бюллетень экспериментальной биологии и медицины. 2010. Т.150, N 7. C. 31-37.
17. Mudraya IS, Revenko SV, Nesterov AV. Bioimpednce harmonic
analysis as a tool to simultaneously assess circulation
and nervous control. // Physiol.Meas. 2011. Vol. 32.
P. 959-976.
18. Ходырева Л.А., Дударева А.А., Мудрая И.С., Маркосян Т.Г., Ревенко С.В.., Кумачев К.В., Логвинов Л.А.
Оценка эффективности ударно-волновой терапии пациентов с тазовой болью методом гармонического анализа
пенильного биоимпеданса. // Бюллетень экспериментальной биологии и медицины. 2013. N 2. С. 259-264.
19. Cheng CL1, de Groat WC. Effect of ovariectomy on external
urethral sphincter activity in anesthetized female rats.
// J Urol. 2011. Vol. 186, N 1. P. 334–340.
20. Halter RJ, Hartov A, Heaney JA, Paulsen KD, Schned AR.
Electrical impedance spectroscopy of prostatic tissues. //
IEEE Trans Biomed Eng. 2007. Vol. 54, N 7.
P. 1321-1327.
21. Keshtkar A. Why is the measured impedance of the bladder
tissue different from the computational modeling results?
// IFMBE Proceedings. 2007. Vol.17. P. 28-31.
22. Giuliano F, Ückert S, Maggi M, Birder L, Kissel J, Viktrup
L. The mechanism of action of phosphodiesterase type 5 inhibitors
in the treatment of lower urinary tract symptoms related
to benign prostatic hyperplasia. // Eur Urol. 2013. Vol.
63, N 3. P. 506-516.
23. Roosen A, Blake-James BT, Wood D, Fry CH. Clinical and
experimental aspects of Adreno-muscarinic synergy in the
bladder base and prostate. // Neurourol Urodyn. 2009. Vol.
28, N 8. P. 938-43.
24. Meng E, Lin W., Lee WC, Chuang YC. (2012), Pathophysiology
of Overactive Bladder. // LUTS: Lower Urinary Tract
Symptoms. 2012. Vol.4, Issue Supplement s1. P. 48–55. 48–55.
doi: 10.1111/j.1757-5672.2011.00122.
25. White CW, Short JL, Haynes JM, Matsui M, Ventura S.
Contractions of the mouse prostate elicited by acetylcholine
are mediated by M(3) muscarinic receptors. // J Pharmacol
Exp Ther. 2011. Vol. 339, N3. P. 870-877.
Кирпатовский В.И., Надточий О.Н., Мудрая И.С., Кабанова И.В.












Комментарии